Slow CCL2-dependent translocation of biopersistent particles from muscle to brain.
Khan Z1, Combadière C, Authier FJ, Itier V, Lux F, Exley C, Mahrouf-Yorgov M, Decrouy X, Moretto P, Tillement O, Gherardi RK, Cadusseau J.
Abstract
BACKGROUND:
Long-term biodistribution of nanomaterials used in medicine is largely unknown. This is the case for alum, the most widely used vaccine adjuvant, which is a nanocrystalline compound spontaneously forming micron/submicron-sized agglomerates. Although generally well tolerated, alum is occasionally detected within monocyte-lineage cells long after immunization in presumably susceptible individuals with systemic/neurologic manifestations or autoimmune (inflammatory) syndrome induced by adjuvants (ASIA).
METHODS:
On the grounds of preliminary investigations in 252 patients with alum-associated ASIA showing both a selective increase of circulating CCL2, the major monocyte chemoattractant, and a variation in the CCL2 gene, we designed mouse experiments to assess biodistribution of vaccine-derived aluminum and of alum-particle fluorescent surrogates injected in muscle. Aluminum was detected in tissues by Morin stain and particle induced X-ray emission) (PIXE) Both 500 nm fluorescent latex beads and vaccine alum agglomerates-sized nanohybrids (Al-Rho) were used.
RESULTS:
Intramuscular injection of alum-containing vaccine was associated with the appearance of aluminum deposits in distant organs, such as spleen and brain where they were still detected one year after injection. Both fluorescent materials injected into muscle translocated to draining lymph nodes (DLNs) and thereafter were detected associated with phagocytes in blood and spleen. Particles linearly accumulated in the brain up to the six-month endpoint; they were first found in perivascular CD11b+ cells and then in microglia and other neural cells. DLN ablation dramatically reduced the biodistribution. Cerebral translocation was not observed after direct intravenous injection, but significantly increased in mice with chronically altered blood-brain-barrier. Loss/gain-of-function experiments consistently implicated CCL2 in systemic diffusion of Al-Rho particles captured by monocyte-lineage cells and in their subsequent neurodelivery. Stereotactic particle injection pointed out brain retention as a factor of progressive particle accumulation.
CONCLUSION:
Nanomaterials can be transported by monocyte-lineage cells to DLNs, blood and spleen, and, similarly to HIV, may use CCL2-dependent mechanisms to penetrate the brain. This occurs at a very low rate in normal conditions explaining good overall tolerance of alum despite its strong neurotoxic potential. However, continuously escalating doses of this poorly biodegradable adjuvant in the population may become insidiously unsafe, especially in the case of overimmunization or immature/altered blood brain barrier or high constitutive CCL-2 production.

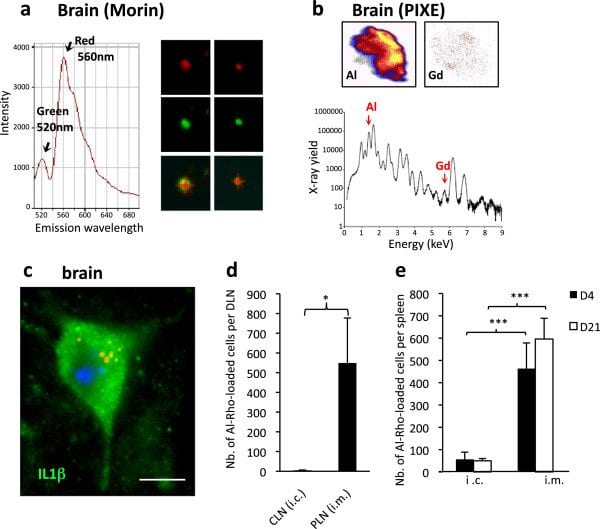
Aluminum deposits in tissues following injection of alum-containing vaccine in the TA muscle. a) Granuloma composed of PAS+ cells is formed in the injected muscle envelope; b) PIXE mapping shows muscle Al deposits in pseudocolors, with confirmatory Al emission spectrum (d21); c) Section of spleen tissue (left panel) displays the large 500 × 500 μm and restricted 100 × 100 μm protonized fields corresponding to the PIXE maps (middle and right panel, respectively) enclosing eligible Al spots (d21); d) Section of brain tissue (left left panel) displays the restricted 100 × 100 μm protonized field corresponding to the PIXE map (middle panel) enclosing eligible Al spot (d21); the number of fields containing one or more Al spots was increased at all tested time points compared to unvaccinated (right panel) mice. (bars: 100 μm). d, day; PIXE, particle induced X-ray emission, TA tibialis anterior.

FLB translocation in DLN following injection in TA muscle. a) Marked translocation of FLBs in parafollicular areas of popliteal DLNs (d4); b) Flow cytometry showing that most FLB-loaded cells extracted from DLN express CD11c at either an intermediate or strong level (d4); c) Immunocytochemistry on CD11b+ cells extracted from DLNs were usually Gr1+/Ly6C+, especially when they had ingested a few particles (left), whereas heavily loaded ones were often Gr1–/Ly6C– (right); d) The number of FLB-loaded cells peaked at d4 post-injection in both popliteal and inguinal DLNs; e) The migration inhibitor BW245C co-injected with FLBs in muscle markedly decreased the number of FLB-loaded cells detected in DLNs at d4 post-injection. The effect was more pronounced in the downstream inguinal DLN; f) The migration inhibitor BW245C co-injected with FLBs in muscle markedly decreased the number of FLB-loaded cells detected in DLNs at d4 post-injection. The effect was more pronounced in the downstream inguinal DLN; (histograms: n = 3 per group, mean + SD, * P <0.05, ** P <0.01, *** P <0.005; bars: 100 μm [a]; 5 μm [c]). d, day; DLNs, draining lymph nodes; FLBs, fluorescent latex beads; TA, tibialis anterior.
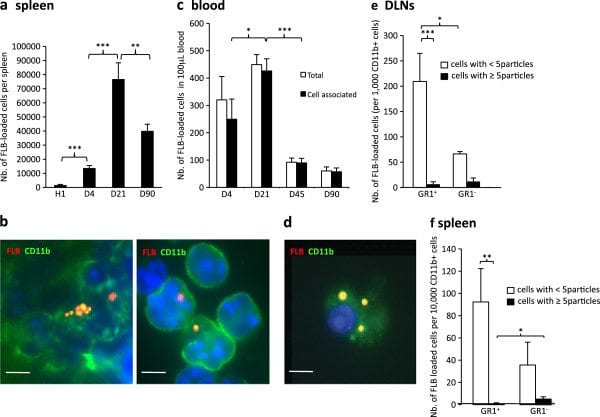
FLB biodistribution in spleen and blood following injection in TA muscle. a) The number of FLB-loaded cells peaked at d21 in spleen; b) In spleen, FLBs were detected in CD11b+ cells as assessed by immunohistochemistry (left) or after cell sorting (right); c) On blood smears, most FLBs were cell-associated from d4 and peaked at d21 post injection; circulating FLB-loaded cells were still detected at d90 endpoint; d) Circulating FLB-loaded cells were CD11b+ (d21); e,f) GR1/Ly6C immunophenotyping of CD11b+ cells that have ingested FLBs. Most are Gr1+/Ly6C+ both in DLNs at d4 (a) and in spleen at d21 (b). (histograms n = 3 per group, mean ± SD, * P <0.05, ** P <0.01, *** P <0.005; bars: 5 μm). d, day; FLB, fluorescent latex beads; n, number; TA, tibialis anterior.

Brain translocation of FLBs following injection in TA. a) Cerebral translocation of FLBs was delayed but relentless until the d90 endpoint in C57 mice and the d180 endpoint in the CX3CR1GFP/+ mouse; b) Unstained section of the brainstem in a C57 mouse at d21 post-injection showing FLBs mostly distributed in the subpial region; c) FLBs distribution in the brain: areas enriched in FLBs were reported on semi-serial rostro-caudal sections of mouse brain stained by Cresyl violet (A to G), using dots of different colors according to the considered time point (d21 to d365) after i.m. injection. Report was done regardless of the number of enclosed particles in each selected area. Note that FLBs were always predominantly found in the grey matter without prominent accumulations at any specific neuroanatomical site. (histograms: n = 3 per group, mean + SD, * P <0.05, ** P <0.01, *** P <0.005; bar in b: 50 μm). d, day; FLBs, fluorescent latex beads; n, number; TA, tibialis anterior.
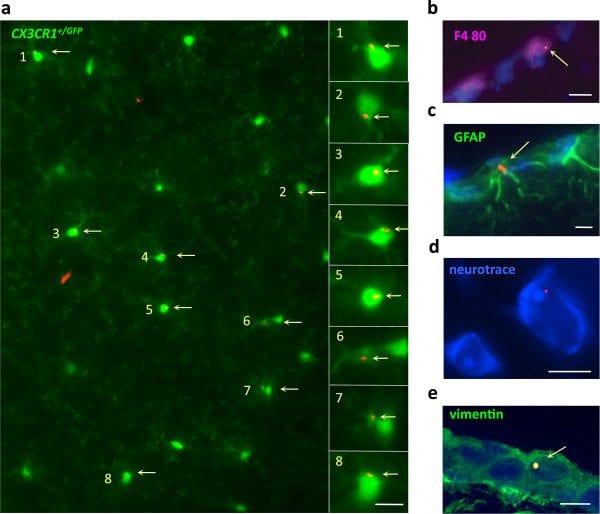
FLBs in various neural cells. a) Unstained section of the brain parenchyma of a CX3CR1GFP/+ mouse at d90 post-injection showing individual FLBs in a significant proportion of GFP+ ramified microglial cells; b-e) In brain of C57 mice at d21post-injection, FLBs were detected in F4/80+ perivascular macrophages (b), GFAP+ astrocytes (c), neurotrace®+ neurons (d), and vimentin+ pial cells (e); (bars: 10 μm). d, day; FLB, fluorescent latex beads.

Mechanisms of FLBs translocation. a) Compared to the i.m. route, direct injection of FLBs in the tail vein of C57 mice was associated with almost no brain translocation at both d21 and d90 post-injection; b) Popliteal and inguinal DLN ablation was associated with a marked decrease of FLB-loaded cells in blood, spleen and brain at d21 post-injection; c) The mdx mouse with altered BBB showed a marked increase of the perivascular CD11b+ cell population, and significant angiogenesis assessed by an increase of CD31+ endothelial cells, compared to normal C57 mice; d–e)Mdx mice showed increased incorporation of FLBs in brain; compared to C57 mice, mdx mice had increased FLB neurodelivery at both d21 and d90, as assessed by both histology (d) or after CD11b+ cell sorting (e); f) At d21, FLBs were mainly detected outside capillary basement membranes immunostained for collagenIV (upper panel), closely associated with CD11b+ perivascular macrophages (lower panel); (histograms: n = 3 per group, mean + SD, * P <0.05, ** P <0.01, *** P <0.005; bar in d: 10 μm). d, day; DLN, draining lymph nodes; FLB, fluorescent latex beads; n, number.
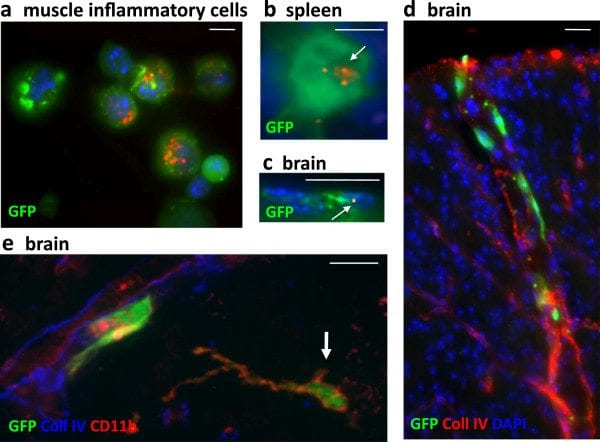
GFP+BM chimeric mice. a-c) Chimeric mice injected intramuscularly with FLBs showed GFP+ BM-derived cells enclosing FLBs among inflammatory cells extracted from the injected muscle (a) at d4 after FLB injection, in spleen (b) and brain (c) at d33 after FLB injection. d-e) Chimeric mice showed incorporation of GFP+ cells in the brain, mainly in the form of perivascular cells in the cortex (d) and, occasionally, in more deeply located ramified CD11b+ cells (e, arrow) at d180 post-BM transplantation. (bars: 10 μm). BM, bone marrow; d, day; FLB, fluorescent latex beads.
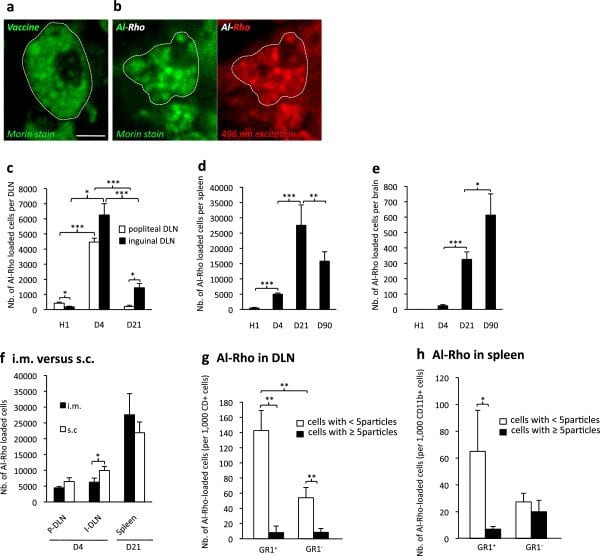
Biodistribution of Al particles. a) Morin stain for aluminum shows rounded alum cytoplasmic agglomerates within muscle macrophages after i.m. vaccine administration in C57 mouse; b) Morin stain confirms that phagocytosed Al-Rho nanohybrids are associated with Al and form particles similar in size to alum agglomerates; c-e) Al-Rho nanohybrids show a time-dependent distribution in DLNs, spleen, and brain strikingly similar to that of FLBs; f) Al-Rho injected by the s.c. route translocate to DLNs and spleen, as observed with the i.m. route; g,h) Ly6C immunophenotyping of CD11b+ cells that have ingested Al-Rho: most are Gr1+/Ly6C+ both in DLNs at d4 (g) and spleen at d21 (h). (histograms: n = 3 per group, mean ± SD, * P <0.05, ** P <0.01, *** P <0.005; bar in a: 10 μm). d, day; DLN, draining lymph nodes; FLBs, fluorescent latex beads; n, number.

CCL2-dependent systemic translocation of Al-particles. a) CCL2 deficient mice show a dramatic decrease of Al-Rho translocation from the injected muscle to inguinal DLN, blood, spleen and brain, as compared to their respective controls (100%). Note that the difference is significant but less pronounced for popliteal DLN; b) rCCL-2 co-injection with Al-Rho is associated with a marked increase of Al-Rho translocation from the injected muscle to inguinal DLN, blood, spleen and brain, compared to their respective controls (100%). Note that the difference is significant but less pronounced for popliteal DLN; c) rCCL-2 infused by an osmotic micropump into the striatum for 15 days is associated with a significant increase of Al-Rho translocation from the injected muscle to brain; d) Combined i.m. and i.c. injection of rCCL2 is associated with a dramatic increase of FLB translocation from muscle to both blood and brain; e) Alum-containing vaccine injected into muscle of CCL-2-deficient, normal, and rCCL-2 mice was associated with a trend of CCL-2-dependent increase of Al concentration levels in brain; (histograms: n = 3 per group, mean ± SD, * P <0.05, ** P <0.01, *** P <0.005, except [e]: n = 10 per group, mean ± SEM). Al-Rho, Al(OH)3 rhodamine nanohybrid; DLN, draining lymph nodes; FLB, fluorescent latex beads; n, number; SEM, standard error of the mean.